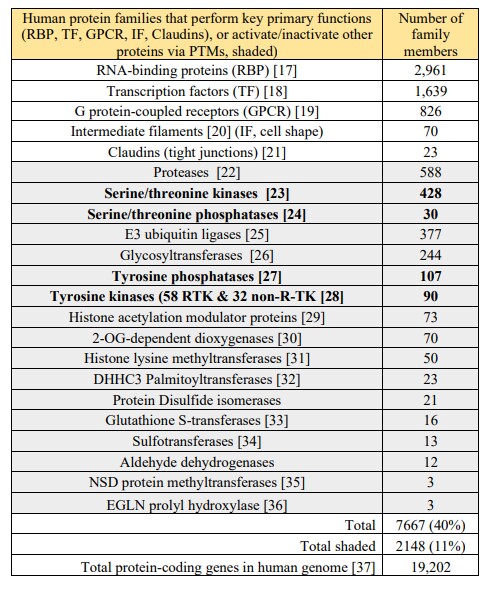
The fact that the serine-threonine kinase/phosphatase ratio (428/30) is so high suggests that this post-translational modification (PTM) is common and mostly irreversible. This hypothesis is
supported by the fact that serine/threonine phosphorylation is the most abundant PTM of the human proteome. About 13,000 (68%) of the ~19,000 proteins in the human genome are
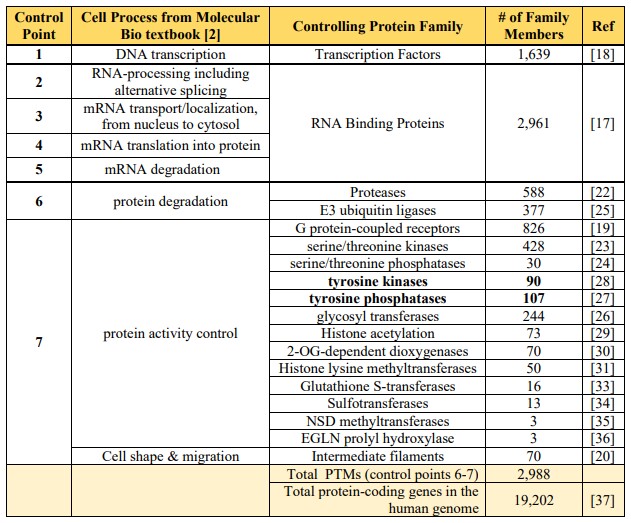
phosphorylated on either serine or threonine residues [38]. In the textbook “Molecular Biology of the Cell” (7th edition, p 401), Alberts et al. discuss seven control points for pathways that lead to cellular differentiation [2] Fig. 7-8. The fact that the PTM protein families in Table 1 match well to control points described by Alberts et al. (Table 2) suggests that PTM timing is key to cell differentiation. Table 2. Correlation of cell processes described in the textbook by Alberts et. al. [2] by protein PTM families comprising 40% of the human genome. A huge family of 1639 transcription factors bind DNA to stimulate or inhibit gene transcription (control point 1). An even larger family of 2961 RNA binding proteins regulates mRNA alternative splicing (control points 2-5). PTMs, including proteases (588 members) and ubiquitin ligases (377 members) regulate protein degradation (control point 6). A myriad (428) of kinases regulate serine/threonine phosphorylation while 90 tyrosine kinases and 107 tyrosine phosphatases, often cancer drivers, cell growth.
Leave a Reply
You must be logged in to post a comment.